Tudomány, 1990, VI/6.
A polimeráz-láncreakció felfedezése
DNS-darabok elôállítása korlátlan
mennyiségben, egy meglepôen egyszerû eljárás
révén a gondolat valószínûtlen körülmények
között, a kaliforniai hegyvidéken született meg egy
holdfényes éjszakán
Írta: Kary B. Mullis*
 Az
embernek néha egészen váratlan pillanatokban jut eszébe
valami jó gondolat. Naivitásom, egy-két szerencsés
melléfogásom, meg a véletlenek összjátéka
folytán 1983 áprilisában, egy péntek este támadt
egy ötletem. Éppen Észak-Kalifornia hegyei felé
autóztam a kacskaringós, holdfényes úton, amikor
megfogalmazódott bennem az azóta polimeráz-láncreakcióként
ismertté vált folyamat gondolata, amelynek lényege,
hogy korlátlan számú génmásolat készítését
teszi lehetôvé.
Az
embernek néha egészen váratlan pillanatokban jut eszébe
valami jó gondolat. Naivitásom, egy-két szerencsés
melléfogásom, meg a véletlenek összjátéka
folytán 1983 áprilisában, egy péntek este támadt
egy ötletem. Éppen Észak-Kalifornia hegyei felé
autóztam a kacskaringós, holdfényes úton, amikor
megfogalmazódott bennem az azóta polimeráz-láncreakcióként
ismertté vált folyamat gondolata, amelynek lényege,
hogy korlátlan számú génmásolat készítését
teszi lehetôvé.
A genetikai információt hordozó DNS-molekula egyetlen
példányából a polimeráz-láncreakció
útján egy délután folyamán százmilliárd
másolatot lehet elôállítani. A reakció
egyszerû: nem kell hozzá más, mint egy kémcsô,
néhány közönséges vegyszer és egy
hôforrás. A másolandó DNS-minta lehet tisztított
anyag, de lehet természetes állapotú, rendkívül
összetett biológiai struktúrák parányi
része is; származhat kórházi szövetmintából,
hajszálból, a bûntett helyén talált beszáradt
vércseppbôl, mumifikálódott agyszövetbôl,
sôt akár jégbe fagyott, negyvenezer éves gyapjas
mamutból is.
Az idõközben eltelt hét év alatt a polimeráz-láncreakció
a biológiai tudományok nagyon sok területén polgárjogot
nyert: különféle alkalmazásairól eddig több
mint ezer közlemény jelent meg. Tulajdonképpen rejtély,
hogy miért nem elôbb derült fény erre a tudományos
szempontból nagy jelentôségû, ugyanakkor egyszerûen
kivitelezhetô folyamatra, hiszen mindaz, ami szükséges
hozzá, már több mint tizenöt évvel korábban
rendelkezésünkre állt.
A polimeráz-láncreakció jócskán megkönnyíti
a molekuláris biológiával foglalkozók dolgát:
alkalmazásával annyit állíthatunk elô
a kívánt DNS-bôl, amennyit csak akarunk. A DNS-molekulát
szokás úgy emlegetni, mintha bármikor elô lehetne
venni a hûtôszekrénybôl, holott a rendkívül
egyszerû szerkezetû vírusokon kívül minden
szervezetbôl nehéz eredeti formában DNS-molekulákat
nyerni.
A nehézség a DNS-molekula természetében
rejlik. Maga a molekula négyféle dezoxi-nukleotidból:
dezoxi-adenilátból (A), dezoxi-timidilátból
(T), dezoxi-guanilátból (G) és dezoxi-citidilátból
(C) álló törékeny lánc, amelyben a genetikai
információt a nukleotidok sorrendje hordozza. Egyszálú
formában a DNS elég ritka; az egymást kiegészítô
bázissorrendû DNS-szálak általában párt
képeznek, és kettôs spirált hoznak létre,
amelyben az egyik szál A nukleotidjai a másik T-jeihez kötôdnek,
a G-k pedig a C-kkel kapcsolódnak. A sejtben a DNS kettôs
spirálját különféle fehérjék
veszik körül; hatásukra a DNS-molekula még inkább
felcsavarodik. A rendkívül hosszú és vékony
DNS-molekulákat igen nehéz tisztán kivonni a sejtbôl;
már csekély nyíróerô hatására
is darabokra törhetnek. Vonjuk ki akár ezernyi egyforma sejtbôl
a DNS-molekulákat: a kívánt génbôl megkapjuk
ugyan mind az ezer példányt, de az egyes példányokat
a véletlenszerûen bekövetkezô törések
folytán más és más DNS-darabok tartalmazzák.
Ez a probléma éveken át nagyon megnehezítette
a gének tanulmányozását. A hetvenes években
azonban felfedezték a restrikciós endonukleázoknak
nevezett enzimeket, amelyek a DNS-szálat meghatározott pontokon
hasítják el. Az endonukleázok segítségével
a DNS-t már kisebb, stabilabb és könnyebben azonosítható
részletekre lehetett darabolni, így a kívánt
gént tartalmazó DNS-darabok kiválogatása is
könnyebbé vált.
A hetvenes évek végén a biokémikusok már
szorgosan vizsgálták a DNS-t az endonukleázok és
az úgynevezett oligonukleotid szondák segítségével.
Az oligonukleotidok rövid, meghatározott nukleotidsorrendû
láncok, amelyek megfelelô körülmények között
specifikusan kötôdnek az egyszálú DNS kiegészítô
(komplementer) nukleotidszekvenciáihoz. A mesterségesen elôállított,
radioaktív jelöléssel ellátott oligonukleotidok
"keresômolekulaként", szondaként használhatók
annak eldöntésére, hogy a vizsgált DNS-mintában
jelen van-e valamely meghatározott gén, illetve nukleotidszakasz.
Ilyen oligonukleotid szondák elôállításával
bízott meg 1979-ben akkori munkaadóm, a Cetus Corporation
(Emeryville, Kalifornia). 1983-ra azonban az izgalmas, de meglehetôsen
veszôdséges "kézi" oligonukleotid-szintetizálás
átadta helyét a kevésbé igézô,
ugyanakkor nagyon megbízható automatizált eljárásnak.
Óriási elôrelépés volt ez, amelyet a
szakmabeliek zöme, köztükjómagam is nagy örömmel
üdvözöltem. Bennünket, nukleotidvegyészeket
azonban ez a kisebbfajta ipari forradalom többé-kevésbé
munkanélkülivé tett. Csak annyi dolgunk maradt, hogy
feltöltsük és ellenôrizzük a laboratóriumi
berendezéseket, amelyek szinte több oligonukleotidot gyártottak,
mint amennyi a mélyhûtõben elfért mindenképpen
többet annál, amennyit a molekuláris biológia
minuciózus tudományának mûvelôi fel tudtak
használni kísérleteikben.
Így azután laboratóriumomban a Cetus Corporationnél
jócskán volt idôm elmélkedni és szöszmötölni.
Egy idô után azon kaptam magam, hogy oligonukleotidokkal pepecselek...
Tudtam, hogy nagy szükség volna egy olyan eljárásra,
amellyel azonosítani lehetne a DNS-molekulák meghatározott
pontján levô nukleotidokat fôként a bonyolult
DNS-molekulák, például az emberi DNS, illetve a kis
mennyiségben rendelkezésre álló DNS-minták
esetében jött volna jól egy ilyen módszer. Felvetôdött
bennem a gondolat: mi lenne, ha a DNS-polimeráz enzimet és
a DNS nukleotidsorrendjének meghatározására
alkalmas, didezoxiszekvenálásnak nevezett eljárás
egyik
változatát használnám erre a célra?
Ötletem kipróbálására ki is találtam
egy egyszerû kísérletet.
A kísérlet megértéséhez elôbb
vegyünk sorba néhány fontos tudnivalót a DNS-molekulával
kapcsolatban. A DNS-molekula egyik végét a kémia szóhasználata
értelmében "háromvesszô (3'-) végnek",
a másik végét "ötvesszô- (5'`-) végnek"
nevezzük. A DNS kettôs spiráljában a két
szál egymással ellentétes irányban fut: az
egyik szál háromvesszô-vége a másik szál
ötvesszô-végével áll párba, és
viszont.
1955-ben Arthur Kornberg és munkatársai (Stanford Egyetem)
felfedezték a késôbb DNS-polimerázoknak elnevezett
sejtbeli enzimek egyikét. A DNS-polimerázok a sejtben többrendbeli
feladatot látnak el; többek között részt vesznek
a DNS-molekula kijavításában és másolásában.
A különféle DNS-polimerázok tovább tudnak
építeni bizonyos oligonukleotidokat, az úgynevezett
"primereket" (indítószekvenciákat): új meg
új nukleotidokat kapcsolnak azok háromvesszô-végéhez,
de csak akkor, ha a primer elôzôleg hibridizálódik,
azaz hozzákötôdik a szerkezetében neki megfelelô
kiegészítô szál: a "templátként"
(mintaként) szolgáló DNS-szál végéhez.
Az oldatban természetesen jelen kell lenniük a DNS-molekula
építôköveinek, a nukleozid-trifoszfátoknak.
A nukleotid, amelyet a polimeráz beépít, mindig
a mintaként szolgáló templátszál soron
következô bázisának kiegészítôje
(komplementere). Ha a templáton mondjuk adenin (A) következik,
akkor az új szálba timin (T) épül be; a guaninnal
(G) szemben az enzim citozint (C) iktat a születô DNS-láncba.
Ha semmi nem jön közbe, a polimeráz a folyamat ismétlésével
a primer háromvesszô-végétôl eljut egészen
a templát ötvesszô-végéig. A DNS kettôs
spiráljának mindkét szála templátként
mûködik a másik szál lemásolása
vagy kijavítása során.
Most pedig néhány szót a didezoxiszekvenálásról,
amelyet egyik felfedezôje, Frederick Sanger (British Medical Research
Council, Molekuláris Biológiai Laboratórium) után
Sanger-eljárás néven is emlegetnek. A módszer
alkalmas a DNS nukleotidsorrendjének meghatározására,
amennyiben rendelkezésre áll a DNS-polimeráz, a megfelelô
templátok és primerek, valamint az egyes nukleozid-trifoszfátok
és speciálisan módosított változataik,
a didezoxi-nukleozid-trifoszfátok (ddNTP-k). A polimerázok
a közönséges nukleotidokhoz hasonlóan a ddNTP-ket
is hozzákapcsolják a növekvô lánchoz; a
ddNTP-k azonban "pontot tesznek" a lánc végére ott,
ahova bekapcsolódnak, mert megakadályozzák a további
bázisok beépülését. A Sanger-eljárással
így különféle hosszúságú,
ddNTP-vel végzôdô DNS-darabokat kapunk. A felhasznált
ddNTP-k ismeretében, a kapott láncokat hosszúság
szerint sorba rendezve meghatározhatjuk a templátszál
bázissorrendjét. Ha az új szálba például
didezoxi-adenin (ddA) épült be egy bizonyos ponton, a templát
megfelelô pontján nyilvánvalóan az adenin párja,
a timin (T) foglal helyet; didezoxi-guanin (ddG) beépülése
pedig a templátban citozin (C) jelenlétére utal.
Az eljárás általam kiötlött, módosított
változatában csak polimerázra, templátra és
primerekre volt szükség; a nukleozid-trifoszfátokat
kihagytam a reakcióelegybôl. Így okoskodtam: a lánc
növekedése azonnal befejezôdik, amint a végéhez
egy ddNTP kapcsolódik; ha tudom, milyen ddNTP kötôdött
a primerhez, máris azonosíthatom a templátszál
megfelelô, a ddNTP bekötôdési helyével szemközti
bázisát.
A DNS-SZÁLAKAT összekapcsolódott nukleotidok: dezoxi-adenilát
(A). dezoxi-timidilát (T), dezoxi-guanilát (G) és
dezoxi-citidilát (C) sorozata alkotja. A két szál
nukleotidsorrendje komplementer, azaz kiegészíti egymást,
mert az A-val szemben mindig T, a G-vel szemben pedig mindig C foglal helyet
a láncban. Ez a komplementer szerkezet tartja össze a két
szálat. Mindkét szálnak van egy "háromvesszô-
(3'-) vége" és egy "ötvesszô- (5'-) vége",
amelyek egymáshoz képest ellentétes helyzetûek,
ezért antiparallelnek mondjuk ôket.
Akkor még nem tudtam, hogy szekvenálási eljárásom
több okból sem mûködne. A fô problémát
az okozza, hogy az oligonukleotidok néha nemcsak az általunk
kiszemelt DNS-szakasszal hibridizálódnak, és így
elkerülhetetlenül összezavarják az eredményeket.
Az emberi DNS például már olyan hosszú, hogy
még a leggondosabb hibridizálással sem lehet elegendô
pontossággal hozzákötni az oligonukleotidokat egy-egy
kiszemelt részletéhez, így bázissorrendjérôl
ezzel a módszerrel aligha tudnánk meg bármi érdemlegest.
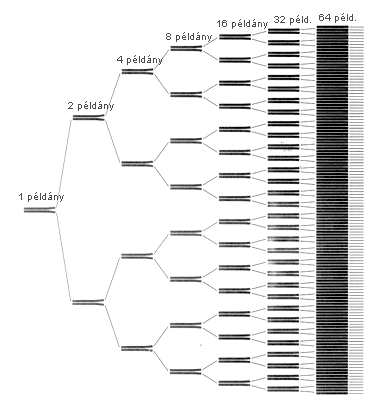
A POLIMERÁZ-LÁNCREAKCIÓ egyszerû eljárás,
a szükséges vegyszerek pedig könnyen beszerezhetôk.
A folyamat DNS-darabok laboratóriumi sokszorosítására
alkalmas. A másolatokból alig néhány óra
alatt már több mint százmilliárd példány
jön létre.
Az emberi DNS-t vizsgáló kutatóknak körmönfontabb
módszerekhez kell folyamadniuk. A DNS-mintát restrikciós
enzimek segítségével különféle hosszúságú
darabokra lehet szabdalni, és ezeket a kisebb DNS-darabokat azután
elektroforézissel el lehet különíteni egymástól.
Ezen az úton a vizsgálandó minta bizonyos mértékig
"megtisztítható" az adott esetben érdektelen DNS-daraboktól.
Ám még így is csak roppant nehezen értékelhetô
adatokhoz juthatunk, ráadásul az eljárás hosszadalmas,
és ha a DNS-minta sérült vagy denaturálódott,
egyáltalán nem alkalmazható. Egy másik lehetôség
a klónozás, amely azonban szintén idôigényesebb
annál, semhogy a rutinszerû DNS-analízist lehetôvé
tenné. Az emberi DNS vizsgálandó szakaszát
a klónozás során egy plazmidnak nevezett kicsiny DNS-gyûrûbe
"varrják bele". A plazmidot és vele a vizsgálandó
DNS-t baktériumokban sokszorosítják, majd oligonukleotid-hibridizációval
és didezoxi-szekvenálással meghatározzák
bázissorrendjét. A nyolcvanas évek elején az
emberi DNS különféle részeinek genetikai információtartalmát
többnyire klónozott DNS-minták didezoxi-szekvenálása
útján sikerült meghatározni.
Kísérletem megtervezésekor botor módon magától
értetôdônek tekintettem, hogy egy emberi DNS-szakaszt
egyetlen oligonukleotid hibridizációjával, mindenfajta
klónozás vagy egyéb trükk híján
is azonosíthatunk. Mentségemül szolgálhat, hogy
Henry Erlich, a Cetus Corporation egyik tekintélyes kutatója
munkacsoportjával abban az idôben szintén egy olyan
eljárással próbálkozott, amely emberi DNS-szakaszok
egyetlen oligonukleotiddal való hibridizációján
alapult.
Egy májusi péntek estén éppen Mendocino
felé autóztam; minden hétvégén felmegyek
ugyanis északra, a vityillómba. Szeretek éjszaka vezetni,
bár a 101-es út nem igényel sok figyelmet. A háromórás
út alatt a kezem dolgozik, a gondolataim viszont szabadon kalandozhatnak.
Azon a bizonyos éjszakán a DNS-szekvenálásról
elmélkedtem.
Elképzelésem egyszerû volt. Elôször melegítéssel
elkülönítem a vizsgálandó DNS két
szálát, majd egy oligonukleotidot hibridizáltatok
az egyik szál megfelelô szakaszával. A DNS-elegyet
négy kémcsôbe osztom szét; mind a négy
csôben jelen van a négyféle ddNTP, csak éppen
az egyik csôben az egyik, a másikban a másik hordoz
radioaktív jelölést. Ezután DNS-polimerázt
adok az elegyekhez, amely minden kémcsôben egyetlen ddNTP-vel
meghosszabbítja a hibridizálódott oligonukleotidot.
A következô lépésben elektroforézissel
elkülönítem a meghosszabbodott oligonukleotidokat a maradék
ddNTP-ktôl, aztán megvizsgálom, a négy közül
melyik radioaktív ddNTP épült be az oligonukleotidokba;
ebbôl már azonosítani tudom a vizsgált szál
megfelelô, az illetô ddNTP-vel szemben beépült
bázisát. Egyszerû, nem igaz?
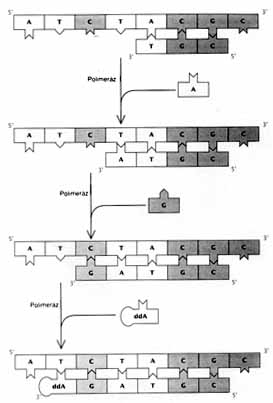
A DNS-POLIMERÁZ nevti enzim képes bizonyos rövid
DNS-szakaszok, úgynevezett oligonukleotid primerek meghosszabbítására,
amennyiben azok valamilyen templáthoz, azaz hosszabb DNS-szakaszhoz
kapcsolódnak. Az enzim a templát nukleotidjait kiegészítô
(azokkal komplementer) nukleotidokat köt a primer háromvesszô-végéhez.
Ha a nukleozid-trifoszfátok didezoxi-változatát, például
didezoxi-adenozin-trifoszfátot (ddA) adunk az oldathoz, a primer
nem hosszabbodhat tovább, mert a láncba beépülô
didezoxi-adenilát (ddA) háromvesszô-végéhez
szerkezeti okokból nem kapcsolódhat további nukleotid.
Cloverdale táján, ahol a 128-as út a 101-esbôl
északnyugat felé ágazik, majd felkanyarodik a hegyek
közé, eszembe ötlött, hogy módszerem egyértelmûbb
eredménnyel szolgálhat, ha kétszálú
DNS-t, és egyetlen oligonukleotid helyett mindjárt kettôt
használok, olyképpen, hogy a két oligonukleotid primer
háromvesszô-vége éppen közrefogja az azonosítani
kívánt bázispárt. Amennyiben a két oligonukleotid
eltérô méretû, meg is tudom különböztetni
ôket egymástól; ha pedig az egyik oligonukleotidot
a DNS-minta egyik szálához, a másikat pedig a másik
szálhoz kötöm hozzá, a két szál bázissorrendjérôl
egyidejûleg nyerhetek adatokat. A kísérlet így
önnön kontrolljául is szolgál, és jócskán
leegyszerûsödik.
Akkor még nem tudtam, hogy ezzel az ötletemmel tehát
azzal, hogy a vizsgált DNS-szakasz két szemközti szálához
háromvesszô-végükkel egymás felé
mutató oligonukleotidokat kötök már közel
jártam a polimeráz-láncreakció felfedezéséhez.
Mindössze arra ébredtem rá hirtelen, hogy még
egy pillanat, és az útmenti árokban találom
magam. . .
Az éjszakai levégô párával és
a virágzó vadgesztenye illatával volt teli. Az út
mellett egyre-másra fehér virágok dugták fejüket
a fényszóró csóvájába. Miközben
a "birtokomon" ásandó kis tavacskákon törtem
a fejem, nem hagyott nyugodni a kérdés, hogy elvben megzavarhatja-e
valami szépen eltervezett bázisszekvenálási
kísérletemet.
Aspiránsként annak idején a Kaliforniai Egyetemen
(San Francisco), Wolfgang Sadee laboratóriumában dolgoztam,
méghozzá többek között John Maybaum társaságában,
aki klinikai célú nukleotidkimutatási módszerek
kidolgozásával foglalkozott. Még tõle értesültem
arról, hogy a DNS-minták nyomokban tartalmazhatnak nukleozid-trifoszfátokat.
Ez bizony tervbe vett kísérletemben jócskán
megnehezítheti az elektroforézis értékelését,
gondoltam, hiszen a mintával bevitt nukleotidok már a jelölt
ddNTP-k hozzáadása elôtt hozzákapcsolódhatnak
a primerek háromvesszô-végéhez.
Mi lenne, tettem fel magamban a kérdést, ha a minta szabad
nukleozid-trifoszfátjait egy bakteriális enzimmel, az alkalikus
foszfatázzal szabdalnám szét? Az enzim leválasztja
a nukleozid-trifoszfátok reakcióképes foszfátcsoportjait,
és ezzel megnyitja az utat a polimeráz-reakció elôtt.
A mintát viszont ezután meg kell tisztítani a foszfatáztól,
mielôtt az a ddNTP-ket is "megemésztené". A nemkívánatos
enzimeket általában melegítéssel lehet hatástalanítani,
mert hô hatására megváltozik a térszerkezetük,
és így mûködésképtelenné
válnak. Úgy gondoltam azonban, hogy a baktériumok
alkalikus foszfatáza visszanyerheti eredeti formáját,
tehát nem alkalmas a probléma megoldására.
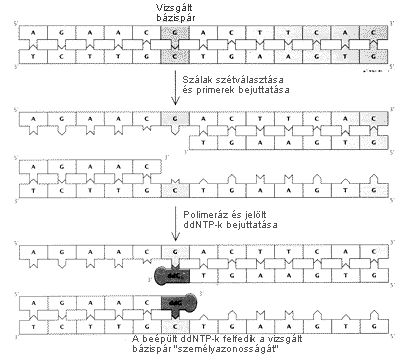
EGY DNS-DARAB valamely bázispárjának azonosítására
a szerzô a didezoxi-szekvenálásnak nevezett eljárás
egy módosított változatát dolgozta ki. Elôször
egy-egy primer kötôdik a DNS két szembenálló
szálához, közrefogva a vizsgálandó bázispárt.
Ezután DNS-polimerázt és didezoxi-nukleozid-trifoszfátokat
(ddNTP) adunk az oldathoz. Így a primerek csak egyetlen bázissal
hosszabbodhatnák meg. A lánchoz kapcsolódott ddNTP-k
azonosításával megtudhatjuk, mi volt a kérdéses
bázis. A kísérlet egyetlen primerrel is mûködik,
de két primer egyidejû alkalmazása az eredmény
ellenôrzésére is módot ad. Így jutott
el a szerzô a polimeráz-láncreakció felfedezéséhez.
Mint kiderült, tévedtem. Csak jóval késôbb
tudtam meg, hogy amennyiben nincsenek cinkionok az oldatban, az alkalikus
foszfatáz hô hatására visszavonhatatlanul "elromlik"
(denaturálódik). Tudatlanságom azonban mégis
áldásnak bizonyult, mert ha ismereteim pontosabbak
lettek volna, aligha nézek jobb megoldás után.
Ahogy haladtam úticélom felé, mérföldrôl
mérföldre újabb meg újabb lehetôségek
jutottak eszembe és bizonyultak használhatatlannak. Közvetlenül
az Anderson-völgy elôtt azonban támadt egy ötletem,
amelyet takarékossági és esztétikai szempontból
is tetszetôsnek találtam. Arra gondoliam, hogy ugyannzt az
enzimet, a DNS-polimerázt kétszer is használhatnám:
elôször a felesleges nukleozid-trifoszfátok eltávolítására,
aztán a jelzett ddNTP-k beépítésére.
Így okoskodtam: ha a minta nukleotidtartalma elegendô ahhoz,
hogy megzavarja a kísérletet, akkor ahhoz is elegendônek
kell lennie, hogy a DNS-polimeráz számára építôanyagot
szolgáltasson. Ha a ddNTP-k hozzáadása nélkül
egy elôkészítô reakciót viszek végbe
az oligonukleotid primerekkel és a polimerázzal, az elegyben
található nukleotidok gyorsan "elfogynak", mert rögtön
az oligonukleotidokhoz kapcsolódnak. A minta hevítésével
azután a meghoszszabbodott oligonukleotidokat különválaszthatom
a vizsgálandó DNS-tôl. Igaz ugyan, hogy e hosszabb
oligonukleotidok továbbra is az oldatban maradnak, ám náluk
sokkal nagyobb számban lesznek jelen a meg nem hosszabbodott primerek,
így az elegy lehûlésekor a vizsgálandó
DNS valószínûleg ezekkel a primerekkel hibridizálódik.
Ezután hozzáadhatom az elegyhez a ddNTP-ket és még
némi polimerázt, s a szekvenálási kísérlet
immár akadálytalanul végbemehet.
Néhány kérdés azonban továbbra is
nyugtalanított. Vajon az elôkészítô reakció
során meghosszabbodott oligonukleotidok nem zavarják-e meg
a késôbbi szekvenálási reakciót? Mi történik,
ha az oligonukleotidokhoz nem pusztán egy vagy kettô, hanem
sok bázis épül hozzá? Mi lesz, ha a meghoszszabbodás
során olyan szekvencia jön létre, amelyen megkötôdhet
a másik primermolekula? Ez bizony gondot jelentene. . .
De nem! Szó sincs róla! Egészen elállt a
lélegzetem a felismeréstôl: a vizsgálandó
DNS-szálaknak és a meghosszabbodott oligonukleotidoknak azonos
lesz a bázissorrendjük! Az elôkészítô
reakció tulajdonképpen megkettôzi a mintában
a vizsgálandó DNS-molekulák számát!
Hirtelen úgy tûnt, hogy a vadgesztenye illata exponenciális
ütemben gyengül. . .
Más körülmények között talán
nem ismertem volna fel ilyen hamar e megkettôzôdés jelentôségét
ugyan mi érdekes lehet egy unos-untalan ismétlôdô
folyamatban? Ám korábban sok idôt töltöttem
számítógépes programok készítésével,
közelebbrôl olyan ciklusokkal, amelyek valamilyen matematikai
mûveletet ismételnek folyamatosan a megelôzô ciklusok
eredményével. Tapasztalatból tudtam tehát,
hogy milyen erôteljes ütemû az egymást követô
megkettôzôdésekbôl adódó exponenciális
növekedés; márpedig a DNS-másolási eljárás,
amelyet kiötlöttem, éppen ilyen ütemû növekedést
eredményezett.
Izgatottan kezdtem sorba venni a 2 hatványait: 2, 4, 8, 16,
32... Homályosan emlékeztem, hogy a 2 tizedik hatványa
körülbelül ezer, vagyis a huszadik hatványa egymillió
körül lehet. Az Anderson-völgy elôtti útelágazásnál
leállítottam a kocsit, és a kesztyûtartóból
papírt-ceruzát vettem elô; sürgôsen ellenôriznem
kellett számításaimat! Utasom bágyadtan érdeklôdött,
hogy miért álltunk meg, és minek gyújtottam
fel a világítást, mire én izgatottan bizonygatni
kezdtem, hogy valami fantasztikus dolgot fedeztem fel. Nem volt különösebb
sikerem; ô újra elszenderedett, én pedig, miután
meggyôzôdtem róla, hogy a 2 huszadik hatványa
valóban egymillió fölötti érték,
továbbhajtottam.
Körülbelül egy mérfölddel arrébb megint
rájöttem valamire. Miután a primerek meghosszabbodása,
a meghoszszabbodott termékek disszociációja, az új
primerek megkötôdése és meghoszszabbodása
néhányszor végbemegy, az exponenciálisan halmozódó
DNS-szálak hossza egyszer csak nem nô tovább, mert
végpontjuk helyét szigorúan meghatárorza, hogy
hova esik a megkötôdött oligonukleotid primerek ötvesszô-vége.
Az eredeti DNS-minta nagyobb darabjait tehát úgy lehetne
sokszorosítani, hogy olyan primereket készítünk,
amelyek egymástól távolabb hibridizálódnak
a DNS-sel. A kapott darabok mindig meghatározott hosszúságú,
jól elkülöníthetô egységek lennének.
Újból leállítottam az autót, és
elkezdtem hibridizálódó, majd meghosszabbodó
DNS-molekulákat rajzolni; a láncreakcióban minden
ciklus terméke mintául szolgált a következô
ciklus számára. Útitársam félálomban
újból tiltakozott. "Nem fogod elhinni! Megáll az ember
esze!" ujjongtam; ô azonban nem osztotta lelkesedésemet...
Házikómat már további megállók
nélkül értük el. Nehéz álmom volt
azon az éjszakán: a fejemben minduntalan "dezoxi-ribonukleáris
bombák" robbantak. . .
Reggel túl fáradtan ébredtem ahhoz, hogy el tudjam
hessegetni a gondolatot: valaki, valahol már bizonyosan kipróbálta
ötletemet. Kutatók ezrei foglalkoznak egyszálú
oligonukleotidok polimerázokkal való meghosszabbításával;
biztos, hogy valamelyiküknek már eszébe jutott a polimeráz-láncreakció.
Ámbár ha valaki már megvalósította volna,
biztosan publikálta volna az eredményeit, és a reakciót
már régóta alkalmazná mindenki a DNS-darabok
sokszorosítására.
Hétfôn bementem a laboratóriumba, és megkértem
George McGregort, az egyik könyvtárost, hogy fussa át
a számítógépben tárolt irodalmi adatokat
a "DNS-polimeráz" címszó szerint. Semmi érdemlegest
nem talált. A következô hetekben megírtam ötletemet
mindenkinek, akit érdekelhetett; a címzettek egyike sem hallott
róla, hogy bárki is próbálkozott volna ilyesmivel,
egyikük sem látott okot arra, hogy ötletem ne lenne megvalósítható,
ugyanakkor egyikük sem lelkesedett érte különösebben.
A DNS-rôl alkotott elképzeléseimet mindig is sokan
tartották fantazmagóriáknak, és nemegyszer
megesett, hogy igazat kellett adnom nekik; most azonban úgy éreztem,
hogy a nyomában vagyok valaminek. . .
Hónapok teltek el, míg elôkészítettem
elsô kísérletemet, kipróbálandó,
hogy a polimeráz-láncreakció mûködik-e.
Ki kellett okoskodnom, hogy milyen pufferoldatot használjak, milyen
legyen a reagensek egymáshoz viszonyított és abszolút
koncentrációja, mennyire melegítsem fel és
hûtsem le az elegyet, mennyi ideig tartson a reakció és
így tovább. Kornbergnek a DNS-polimerázról
írott elsô közleményeibôl indultam ki. A
kísérlethez egy 25 bázispárból álló,
plazmidba zárt DNS-darabot és két, 11, illetve 13
bázisból álló oligonukleotid primert választottam.
Amikor minden készen állt, elindítottam kísérletemet,
mégpedig úgy, ahogy a legjobban szeretek kísérletezni:
adva volt egyetlen kémcsô, a végeredmény pedig
csakis "igen" vagy "nem" lehetett. Megsokszorozza-e a láncreakció
a kiszemelt DNS-szekvenciát? Mint kiderült: megszokszorozza!
Amikor késô este hazaindultam, észrevettem, hogy
Al Halluin, a Cetus szabadalmi ügyvivôje még a szobájában
van. Bementem, és beszámoltam neki felfedezésemrôl,
lefestettem elôtte a polimeráz-láncreakciót.
A körülbelül száz ember közül, akinek eladdig
hírt adtam a dologról, Al volt az elsô, aki jelentôséget
tulajdonított neki, és azon melegében látni
akarta a kísérleti eredményeket igazoló autoradiogramot.
Vannak, akiket nem gyôz meg egyetlen "kémcsôkísérlet",
Al azonban nem tartozott közéjük. Még szobájában
végighallgatta magyarázatomat, és elismerte, hogy
jól hangzik. Most, a laboratóriumban már izgatott
is lett egy kicsit; javasolta, hogy dolgozzak tovább a kísérleten,
és jelentsem is be a szabadalmat, aztán gratulált
és hazament.
A következô hónapokban egy ifjú és rendkívül
tehetséges matematikus, Fred A. Faloona segítségével
vizsgálgattam
és tökéletesítgettem a polimeráz-láncreakciót.
Frednek ez volt a legelsô biokémiai jellegû munkája.
A sikert sörrel ünnepeltük meg.
További néhány hónap alatt meggyôzôdtünk
róla, hogy a polimeráz-láncrakció a plazmid-DNS
nagyobb darabjainak sokszorosítására is alkalmas.
Végül Henry Erlich laboratóriumából szereztünk
némi emberi DNS-t, és bebizonyítottuk, hogy a láncreakcióval
egyetlen génpéldány kiválasztott darabja is
megsokszorozható.
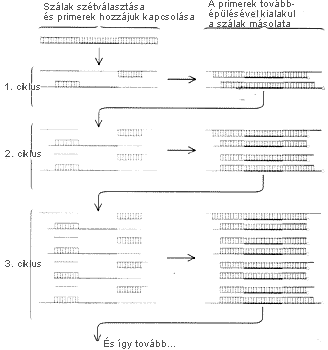
A POLIMERÁZ-LÁNCREAKCIÓ ciklikus folyamat; a DNS-darabok
száma minden ciklusban megkettõzôdik. A vizsgálandó
kétszálú DNS szálait melegítéssel
elkülönítjük, így hûlés közben
primerek tudnak kötôdni hozzájuk. Ezután a DNS-polimeráz
a megfelelô nukleotidok beépítésével
fokozatosan meghosszabbítja a primereket, és végül
létrejönnek az eredeti DNS-szálak pontos másodpéldányai.
A mai napig a polimeráz-láncreakció még
sokat csiszolódott. Több, egymástól némiképp
eltérô változata használatos: jómagam
a DNS-minta 6098 Celsius fokos hômérsékleten való
sokszorosítását tartom a legelônyösebbnek.
A ciklusok ilyenkor csupán egy-két percig tartanak, a vizsgálandó
DNS-molekulák száma pedig minden ciklusban megkettôzôdik.
Az alkalmazott primerek általában 20-30 bázis hosszúságúak.
A legfontosabb hatás, hogy a folyamatot egy hévforrásokban
élô baktérium, a Thermus aqcuaticus DNS-polimerázával
katalizáljuk. A korábban használt polimeráz
ugyanis hô hatására könnyen tönkrement, ezért
minden ciklusban pótolni kellett. A Thermus aquaticus DNS-polimeráza
viszont magas hômérsékleten is stabil és mûködôképes,
így elegendô a reakció kezdetén az elegyhez
adni. Ezt a magas hômérsékleten mûködô
polimerázt ma már génmanipulációval
módosított baktériumokkal termeltetik.
A polimeráz-láncreakcióval elérhetõ,
gyakorlatilag korlátlan génsokszorosítás példa
nélkül állt., ezért a tudományos közvélemény
nem könnyen "emésztette meg". Senki nem számított
arra, hogy létezik olyan folyamat, amellyel mindenféle DNS
korlátlan mennyiségben elôállítható.
Frednek és nekem pofonegyszerûnek tûnt a reakció,
hiszen annyit játszadoztunk vele; a többi kutatónak
azonban idô kellett, hogy hozzászokjon a láncreakció
gondolatához.
Miközben a szabadalmon dolgoztam, 1984-ben egy poszteren is beszámoltam
a polimeráz-láncreakcióról a Cetus Corporation
évente megtartott tudományos ülésén. A
Cetusnál elsôrangú tudósok dolgoznak, így
alig vártam, hogy felfedezésemet megoszthassam velük.
Ám senki nem mutatott érdeklôdést poszterem
iránt: csak ránéztek, aztán továbbmentek
én meg egyre jobban feszengtem. Végül észrevettem
Joshua Lederberget, a Rockefeller Egyetem elnökét, és
odacsaltam, hogy ugyan nézze már meg az eredményeimet.
Josh gondosan végigböngészte a posztert, majd
felém fordította azt a hatalmas, Nobel-díjas fejét,
amelybôl 1946-ban kipattant a baktériumok nemi közösülésének
gondolata. "És ez mûködik is?" kérdezte évôdve.
Örömmel kezdtem magyarázni, hogy igen, mûködik,
aztán még hosszan beszélgettünk. Josh megemlítette,
hogy körülbelül húsz évvel azelôtt,
amikor Kornberg felfedezte a DNS-polimerázt, mindkettejüknek
eszébe jutott, hogy az enzimet valahogyan a DNS sorozatgyártására
is be lehetne fogni, de nem jöttek rá, hogyan. Emlékeztettem
rá, hogy akkoriban még nem volt gyerekjáték
oligonukleotidokhoz jutni, és jóformán egyetlen DNS-szekvencia
sem volt ismert.
Josh ekkor újra a poszterra nézett, ezúttal már
majdnem olyan arckifejezéssel, amilyet vártam tôle.
Azt hiszem, ô volt az elsô, aki a polimeráz-láncreakció
végtelen egyszerûségét látva azt érezte,
ami ma már szinte minden DNS-kutató legelsô gondolata:
"Hogyhogy ez nekem nem jutott eszembe'?!" Fogalmam sincs, miért
éppen bennem fogamzott meg a gondolat egyszerûen belebotlottam
egy májusi éjszakán.
*Kémiai Nobel-díj,1993.